Реакции фоторецепторов на световое излучение


Описание
Суммарный ответ фоторецептора на поток квантов той или иной длины волны света складывается из элементарных дискретных гиперполяризационных реакций, каждая из которых имеет амплитуду 0,2 мВ и возникает на 1 фотон. Отдельный поглолценный фоторецептором квант света с вероятностью 0,7 вызывает фотоизомеризацию молекулы пигмента. В темноте дискретные реакции наблюдаются в наружном сегменте и в отсутствие светового сигнала, что связано со спонтанным (тепловым) распадом молекул фотопигмента.Например, при комнатной температуре в наружном сегменте палочки жабы регистрируется в среднем 1 дискретный ответ за 40—50 с («темновой шум»). Дискретная реакция на 1 фотон сама складывается из большого числа еще более элементарных актов — закрывания отдельных натриевых каналов. По расчетам разных авторов, изомеризация одной молекулы пигмента приводит в результате трансдукции к закрыванию в наружных сегментах палочек от 100 до 300 натриевых каналов, в наружных сегментах колбочек — от 20 до 40.
По оценкам разных авторов, эффект поглощения 1 фотона составляет от 200 мкВ до 6 мВ для палочек и около 100 мкВ для колбочек. Интересно, что вывод о способности отдельной; палочки реагировать на 1 квант света был сделан, по данным психофизических экспериментов, на человеке еще в 1942 г.
При увеличении интенсивности света суперпозиция дискретных ответов образует суммарное отклонение фоторецепторноп потенциала. Закрытие натриевых каналов под влиянием света ведет к возрастанию сопротивления мембраны рецептора. При этом рецептор гиперполяризуется. Амплитуда гиперполяризационного рецепторного потенциала тем больше, чем выше интенсивность света:
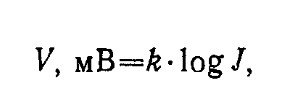
где k — коэффициент пропорциональности, подбираемый так, чтобы V было выражено в мВ. Эта зависимость отражает средний уровень интенсивностей. В зоне слабых раздражений ответ линейно зависит от числа квантов на входе. При очень сильных интенсивностях потенциал достигает плато в области насыщения. Зависимость между амплитудой рецепторного потенциала и интенсивностью света любой длины волны в общем виде описывается уравнением Михаэлиса — Ментон:

где V — амплитуда ответа, I — интенсивность света, Vmах — максимально возможная амплитуда ответа, а — константа, равная интенсивности света, при которой V=Vmax/2; n — показатель, определяющий наклон кривой и варьирующий в зависимости от вида животного и типа фоторецептора. При слабом свете, когда I+?=?, V=VmaxI/?, ответ рецептора линейно связан с интенсивностью.
При больших яркостях света V достигает Vmах («насыщения») и поглощение фотонов перестает быть эффективным. Насыщение ответа происходит при поглощении отдельным фоторецептором всего лишь нескольких сот фотонов (обесцвечивание менее 1% молекул пигмента). Число квантов света, при поглощении которых потенциал темноадаптированного фоторецептора достигает Vmax/2 для разных видов животных (жаба, саламандра, крыса, обезьяна) варьирует в небольших пределах — от 30 до 50.
[banner_centerrs] {banner_centerrs} [/banner_centerrs]
Уравнением Михаэлиса — Ментон описывается также зависимость от интенсивности света натриевой проницаемости наружного сегмента и величины фототока, регистрируемого в условиях фиксации мембранного потенциала фоторецептора. Физико-химические механизмы, лежащие в основе уравнения Михаэлиса — Ментон, применительно к фоторецептору, пока не ясны, но указывают на роль ферментативных реакций.
Принцип унивариантности.
Зависимость амплитуды ответа фоторецептора от интенсивности света (амплитудная функция) имеет одну и ту же форму и наклон для спектральных стимулов разных длин волн в том случае, когда реакция фоторецептора •определяется только одним пигментом.
Это следует из так называемого «принципа унивариантности», сформулированного Нака и Раштоном. Согласно этому принципу фотохимический эффект, вызываемый поглощенным фотоном в наружном сегменте, не зависит от длины волны света. И тогда реакции рецептора на стимулы разного спектрального состава, выравненные по числу поглощаемых (эффективных) фотонов, должны быть идентичны по амплитуде и форме ответа.
Отклонение от принципа унивариантности, выражающееся, в частности, в различиях между амплитудными функциями для излучений разных длин волн, свидетельствует о конвергенции сигналов, передаваемых пигментами с разной спектральной характеристикой. Таким образом, отклонение от принципа унивариантности может служить своего рода тестом на отсутствие или наличие взаимодействия на уровне отдельных фоторецепторов.
Спектральные характеристики рецепторов.
Первые электро-физиологические записи реакций фоторецепторов позвоночных были сделаны японским ученым Томита с сотрудниками в 1965—1967 гг., зарегистрировавшим ответы колбочек сетчатки карпа на равноквантовые монохроматические стимулы разных длин волн. Этот метод принято называть методом «постоянного стимула». Такими постоянными стимулами являются монохроматические излучения, выравненные по числу квантов.
Оказалось, что амплитуды гиперполяризационной реакции отдельной колбочки меняются с изменением длины волны равно-квантового излучения по-разному в зависимости от типа колбочки. Одни колбочки (В-) имеют максимум гиперполяризации при излучении с длиной волны 460 нм, другие (G-) — при 535 нм и третьи (R-) — при 620 нм (рис. 3.1.5а).

В последующих за этим исследованиях было обнаружено, что аналогичными широкополосными функциями спектральных реакций обладают колбочки и палочки у широкого ряда позвоночных (лягушка, черепаха, белка, кошка, обезьяна).
Функция спектральных реакций не совсем точно отражает характеристику спектральной чувствительности фоторецептора так как ее форма зависит от уровня интенсивности равноквантовых излучений, служащих постоянными стимулами. Для построения «кривой спектральной чувствительности» используется метод равных ответов (рис. 3.1.5а).
С этой целью для каждого монохроматического стимула определяется интенсивность излучения, необходимая для того, чтобы вызвать гиперполяризационную реакцию заданной амплитуды (например, «А»), Интенсивность светового стимула Ii принято выражать в логарифмических единицах относительно максимальной величины равноквантового потока света Iо, определяемой мощностью источника световой энергии и свойствами оптической системы фотостимуляции (lg Ii/Io). Затем определяется ?j — длина волны монохроматического стимула, при которой (для вызова фиксированного ответа «А») требуется наименьшее количество энергии стимуляции: lg Ij/Io=min.
Интенсивность каждого монохроматического стимула* вызвавшего такой же по величине ответ «А», нормируется по этому минимуму, и по этим нормированным значениям строится график функции относительной спектральной чувствительности клетки, где по оси абсцисс откладывается длина волны спектрального стимула, а по оси ординат — значения отношения интенсивностей в процентах.
Сравнение функций спектральной чувствительности фоторецепторов со спектрами поглощения содержащихся в них пигментов показало, что в большинстве случаев каждому фоторецептору соответствует свой отдельный фотопигмент. Отличие спектральных характеристик разных фоторецепторов определяется использованием разных пигментов, отличающихся спектрами поглощения. Показательным в этом отношении является соответствие между спектрами поглощения пигментов и функциями спектральных реакций одиночных колбочек в сетчатке рыб (рис. 3.1.5б).
Из анализа формы функции спектральной чувствительности и функции спектральных реакций (рис. 3.1.5а, б) отдельного фоторецептора следует, что для каждого из них в отдельности излучения разного спектрального состава качественно неразличимы. Фотохимический эффект фотона не зависит от его длины волны, которая определяет только вероятность взаимодействия кванта света с молекулой пигмента. Это справедливо в отношении каждой молекулы пигмента независимо от состояния других молекул, например от их распада под действием света.
Отсюда следует аддитивность воздействия излучений на отдельный фоторецептор: если два излучения воздействуют на один и тот же приемник в одно и то же время, то реакция приемника будет равна сумме реакций, вызываемых действием этих же излучений по отдельности. Отсюда же следует постоянство спектра поглощения пигмента и спектральной чувствительности содержащего его фоторецептора при разных уровнях освещения.
Эти фундаментальные свойства фоторецепторов — качественная тождественность эффекта разных излучений и постоянство спектральной чувствительности — лежат в основе «принципа унивариантности». Однако случаются отклонения от этого классического правила. Так, палочки содержат иногда смесь пигментов. Избирательная цветовая адаптация может изменять соотношение пигментов, входящих в смесь, и тем самым изменять спектральную чувствительность фоторецептора.
Другой причиной нарушения стабильности реакций фоторецептора может служить самоэкранирование фотопигмента в наружных сегментах: поверхностно лежащие слои молекул являются нестабильным цветным светофильтром на пути света к более глубоко расположенным молекулам, что отражается на постоянстве спектральной характернее тики фоторецептора.
Электрические контакты между рецепторами.
Электронно-микроскопические и электрофизиологические данные свидетельствуют о том, что соседние фоторецепторы одного типа (G- или R-колбочки, палочки) связаны между собой электрическими синапсами в единую проводящую сеть — синцитий (синцитий G-колбочек, синцитий R-колбочек, синцитий палочек). В силу этого потенциал, генерируемый в одном фоторецепторе, «растекается» по сети связанных с ним рецепторов.
Таким образом, выходная реакция рецептора зависит не только от числа поглощенных квантов света в его собственном наружном сегменте, но к ней добавляется и влияние реакций соседних рецепторов. Для колбочек это влияние распространяется на расстояние до 50 мкм, для палочек — до 250 мкм.
В опытах на некоторых животных обнаружены связи между рецепторами разного типа: между G- и R-колбочками черепахи и даже между палочками и колбочками. «Неунивариантность» спектральных ответов описана также для случая двойных колбочек, основной и дополнительный членики которых могут взаимодействовать посредством близкого смыканияг мембран. Однако такие связи между фоторецепторами разного типа достаточно слабы и выявляются лишь в специальных экспериментальных условиях, минимизирующих взаимодействие между фоторецепторами одинакового спектрального класса.
Усилительное взаимодействие между фоторецепторами разного типа приводит к «смешению цветовых сигналов» и, как следствие, к уменьшению цветового и яркостного контрастов, к «размыванию» границ объекта.
Тормозные взаимодействия между рецепторами.
Если в пределах зоны с диаметром 50-ь70 мкм колбочка усиливает выходную реакцию под влиянием реакции соседних рецепторов, то освещение окружения этой зоны может вызывать некоторую деполяризацию в колбочке, т. е. уменьшать реакцию. Считается, что в основе такого тормозного воздействия между далеко расположенными друг от друга колбочками (внешний диаметр тормозной зоны варьирует в пределах от 200 мкм до 5 мм) лежит система отрицательной обратной связи между колбочками и горизонтальными клетками.
Существует две гипотезы, по-разному объясняющие работу механизма обратной связи. Согласно «химической гипотезе» в темноте с горизонтальных клеток на фоторецепторы поступает гиперполяризующий медиатор (предполагают ГАМК). И тогда гиперполяризация горизонтальной клетки на свету уменьшает выделение тормозного медиатора и вызывает деполяризацию в колбочках, являющуюся, таким образом, уменьшением ТПСП.
Однако эффект обратной связи нестабилен, наблюдается не всегда (в колбочках карпа и карася он, как правило, вообще отсутствует) и быстро исчезает в ходе опыта. Нет до сих пор и определенных морфологических данных о наличии химических синапсов от горизонтальных клеток к колбочкам. Эти трудности обходит «электрическая гипотеза» обратной связи. Данная гипотеза объясняет обратное влияние горизонтальных клеток на рецепторы утечкой электрического тока в самом химическом синапсе, связывающем фоторецептор с горизонтальной клеткой.
Этот ток, генерируемый горизонтальной клеткой под воздействием света, неизбежно затекает «обратно» в синаптическую щель, окружающую инвагинирующий отросток горизонтальной клетки и вызывает падение напряжения, которое фактически оказывается приложенным к пресинаптической мембране рецептора и поэтому влияет на выделение из нее медиатора.
Этот эффект распространяется на любой химический синапс, но здесь он особенно силен, учитывая глубокое врастание отростков этих клеток внутрь синаптических образований рецептора. Сделанная на основании гипотезы физическая модель качественно воспроизводит феноменологию проявления обратной связи как между колбочками и рецепторами, так и в нейронах второго порядка между биполярами и горизонтальными клетками.
Число и типы фоторецепторов, необходимых для различения цветов. Согласно «принципу унивариантности» наличие одного типа фоторецепторов недостаточно для различения цветов: в этом случае излучения любого спектрального состава могут быть уравнены для глаза путем изменения только их энергии. Чтобы цветоразличение было возможно, необходимо как минимум два типа рецепторов с разной спектральной чувствительностью. Тогда излучения с разным спектральным составом будут возбуждать их в разных соотношениях. Анализ этих соотношений осуществляется уже нейронными структурами последующих уровней.
Традиционно цветовое зрение связывают с работой колбочковой (фотопической) системы рецепторов. В сетчатке у животных, с развитым цветовым зрением (человек, обезьяна, черепаха, некоторые рыбы) имеется три типа колбочек с максимумами чувствительности в коротко-, средне- и длинноволновом участках спектра.
Различные типы нарушений цветоразличения сопровождаются редукцией различных типов колбочек: у протанопов («краснослепых») отсутствуют колбочки, содержащие пигмент с максимумом поглощения в длинноволновой области спектра, у дейтеранопов («зеленослепых») — колбочки с средневолновым пигментом. Имеются данные об участии в цветовом зрении также и палочек.
У травяной лягушки Иапа етрогапа в различении коротковолновых излучений участвуют «зеленые» (432 нм) и «розовые» (502 нм) палочки. Причем в диапазоне средних (мезопических) интенсивностей внешнего освещения эти два типа палочек работают совместно с колбочками (575 нм). Имеется ряд исследований, доказывающих участие палочек в цветоразличении и при фотопических уровнях освещенности.
Наличие системы фоторецепторов разного типа является необходимым, но не достаточным условием для цветового зрения. Так, у кошек в сетчатке имеются колбочки и палочки, но, по данным поведенческих опытов, цветовое зрение у них практически отсутствует. В нейрональных структурах должны быть заложены алгоритмы, позволяющие по распределению возбуждений в слое рецепторов определять цвет объекта. У низших позвоночных такие алгоритмы реализуются скорее всего в последующих за фоторецепторами клеточных слоях самой сетчатки. У высших животных системы цветокодирующих нейронов сконцентрированы в структурах наружного коленчатого тела и коры.

Комментариев 0