Моделирование патогенеза начальной близорукости на основе рефракционной биомеханики глаз (Часть 2)


Описание
Необходимо отметить отсутствие в этих схемах достаточной обоснованности объединения хрусталика, цилиарной мышцы и цинновой связки в одну информационно- функциональную единицу, так как сигнал, поступающий от сетчатки к центру управления аккомодацией, проходит через зрительную кору, а ответный сигнал идет уже через цилиарную мышцу.Б. X. Гуревич (1971) предложил схему регулирования движений глазного яблока по ходу направленного поворота к цели с учетом эффектов восприятия, Schultze (1972) — кибернетическую схему окуломоторики, Дж. Милсум (1968) — схему пространственных соотношений в системе зрительной фиксации. Все эти схемы достаточно подробны, но раскрывают сущность лишь конвергенции и сочетанных движений глазных яблок, в то время как в зрительно-моторную цепь обратной связи входят и другие рефлекторные механизмы.
Ряд авторов считает, что стимулом для развития аккомодации является дефокусировка изображения на сетчатке (Fincham, 1953). В последнее время появились работы, в которых учитывается расфокусировка ретинального изображения при перемещении тест-объектов. Так, в формальной модели контура регулирования аккомодации (Toates, 1970, 1972) учтена связь реальной преломляющей силы хрусталика и степени расфокусировки ретинального изображения. Ronchi и Fontana (1971) показали, что устойчивость зрительной системы к дефокусировке изображения выше в случае более сильной аккомодации и меньшего диаметра зрачка.
Значительную роль в фокусировке изображения на сетчатке играют флюктуации аккомодации. Их частота, по Campbell, Robson, Westheimer (1959), равна 0,5 Гц, а амплитуда — 0,2.
Таким образом, приведенные данные говорят о том, что при зрительной работе аккомодационная установка глаз происходит рефлекторно. В отношении возможности непосредственного воздействия на аккомодацию через центральную нервную систему весьма убедительны результаты исследований Chin, Ishikawa, Lappin (1968). При электрической стимуляции центров аккомодации среднего мозга у обезьян они выявили линейную зависимость между величиной стимула и напряжением аккомодации. Ниже мы приводим данные наших исследований (1973, 1975).
Под автофокусировкой понимается способность зрительного анализатора постоянно обеспечивать четкое восприятие объекта, в том числе при его перемещении в пространстве.
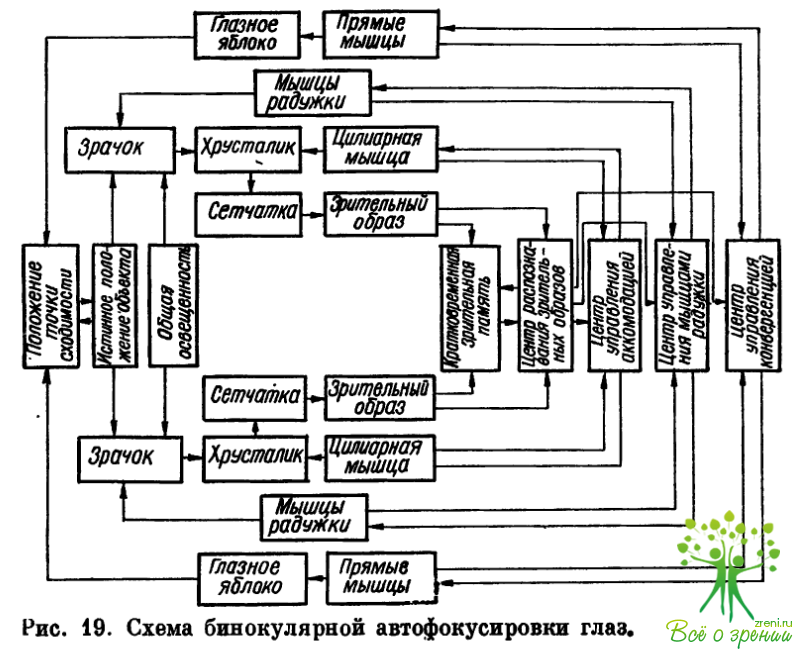
На рис. 19 представлена предложенная нами (1973) схема, иллюстрирующая ход процесса бинокулярной автофокусировки (аккомодационное, конвергентное и зрачковое кольца рефлексов). Стрелки указывают направление сигналов или направленное воздействие (например, цилиарная мышца воздействует на хрусталик).
Опишем кратко механизм действия различных методов лечения и диагностику спазма аккомодации. Пусть рассматриваемый предмет находится на таком расстоянии от глаз, что изображение его на сетчатке четкое. Поставленная перед глазом слабая отрицательная линза изменяет ход параллельных лучей на расходящиеся и отдаляет их фокус кзади от сетчатки, что происходит и при рассматривании близкого предмета неаккомодирующим глазом. Следовательно, такая линза имитирует приближение объекта к глазу, а положительная, наоборот, их отдаление. В обоих случаях изображение на сетчатке станет нечетким.
Центр распознавания зрительных образов может произвести сравнительный анализ изменений, происшедших с изображением, для чего достаточно сравнить нечеткий зрительный образ с имеющимся в кратковременной зрительной памяти четким образом. По изменению размеров изображения центр распознавания получает возможность дать центрам управления аккомодацией и конвергенцией правильную информацию о том, приблизился объект или удалился.
Для того чтобы вновь получить четкое изображение на сетчатке, центр аккомодации «отдает приказ» о расслаблении (опыт с положительными линзами) или сокращении (опыт с отрицательными линзами) цилиарной мышцы, что компенсирует влияние линз.
Прямые экстраокулярные мышцы — наружная и внутренняя — получают сигналы от центра конвергенции. О своем состоянии, как это видно из схемы, цилиарная мышца постоянно «извещает» центр аккомодации, а прямые мышцы — центр конвергенции. В конечном счете положение точки сходимости вновь совпадает с видимым положением объекта, а фокус вновь попадает на сетчатку. Изображение предмета становится четким.
Если сравнить с описанными опытами разработанные А. И. Дашевским методики тренировки аккомодации путем постепенного увеличения силы отрицательных линз и расслабления спазма аккомодации микрозатуманиванием, то становится очевидным, что они, по существу, опираются на принцип автофокусировки глаз. Именно рефлекторная основа этих методов обусловливает их высокую эффективность при лечении ложной миопии.
В настоящее время используют четыре принципиально различных способа воздействия на аккомодацию и конвергенцию, основанные на приведенной выше схеме бинокулярной автофокусировки глаз. Два из них вызывают расслабление спазма аккомодации (призмы основанием к носу и положительные линзы), два других — ее усиление (призмы основанием к виску и отрицательные линзы).
[banner_centerrs] {banner_centerrs} [/banner_centerrs]
Эти способы используют для тренировок как на близкое расстояние, так и вдаль при сохранении их аккомодационно-конвергентной рефлекторной основы.
Разработанные А. И. Дашевским методы лечения ложной миопии основаны на принципе автофокусировки глаз и являются конкретным клиническим воплощением различных способов воздействия на аккомодацию и конвергенцию.
Цилиарная мышца не выполнит «непосильного приказа» центра управления аккомодацией, поэтому величина шага, на который изменяется сила линз при оптическом микрозатуманивании, должна быть небольшой. В противном случае все преимущества рефлекторных тренировок не имеют смысла. С учетом этого в практически используемых методах при тренировке аккомодации шаг обычно равен 1,0 Д, а при лечении микрозатуманиванием +0,25 Д. Однако задача выбора оптимальных дозировок силы линз еще не может считаться решенной окончательно. Поэтому интересна связь между оптимальной величиной шага при микрозатуманивании и глубиной фокусной области человеческого глаза.
Н. М. Сергиенко (1967, 1975) измерял глубину фокусной области различными методами и получил при этом величину порядка 0,5—0,6 Д. Механизм автофокусировки эффективно функционирует при таких перемещениях объекта, которые не выводят его изображение за пределы фокусной области. При микрозатуманивании таким перемещениям соответствует шаг от самого малого до 0,5— 0,6 Д. Практически обоснован выбор шага в 0,25 Д, однако шаг в 0,12 Д оказывается эффективнее. В связи с этим при стойком и полустойком спазме полезно применять дистантный и оптико-дистантный методы, так как в этих случаях шаг может быть равен и 0,01 Д (А. И. Дашевский, Е. И. Кузина, А. А. Ватченко, 1976).
Моделирование аккомодации и ее спазма. Несмотря на наличие оригинальных теорий аккомодации, наиболее обоснованной по-прежнему остается теория Гельмгольца (1855). Считается, что у человека существует и экстраокулярная аккомодация, не связанная с хрусталиком (внешняя аккомодация). Однако главную роль играет хрусталиковая аккомодация. Ее разделяют на внекапсулярную (изменение кривизны поверхностей хрусталика) и внутрикапсулярную (изменение показателя преломления слоев хрусталика), которую детально исследовал Гулыпранд (1912). В основе внекапсулярной и внутрикапсулярной аккомодации лежит один биомеханический процесс, связанный с сокращением цилиарной мышцы и ее воздействием на хрусталик, впервые описанным Гельмгольцем.
Во время покоя аккомодации циннова связка натянута и оказывает некоторое давление на капсулу хрусталика. При сокращении цилиарной мышцы волокно цинновой мышцы расслабляется, силы, действующие на хрусталик, уменьшаются. Благодаря своей эластичности он становится более выпуклым. Расслабление волокон цинновой связки при аккомодации подтверждается феноменом Гесса (1908— 1909), который обнаружил, что хрусталик при аккомодации опускается на 0,25—0,3 мм под давлением собственной массы.
Для построения биомеханической модели аккомодации или конвергенции простейшую расчетную схему глаза необходимо детализировать и приблизить к реальному анатомическому строению глаза. Вполне допустимо первоначально абстрагироваться от деталей, как неравномерность толщины капсулы хрусталика на отдельных участках, неточность представления передней и задней частей хрусталика в виде шаровых сегментов, феномен Гесса, некоторое уменьшение при аккомодации радиуса кривизны в центре передней поверхности хрусталика при одновременном увеличении его на периферии этой поверхности (Brown, 1973).
Только в первом приближении можно использовать расчетную схему глаза, в которой хрусталик представлен оболочкой, заполненной жидкостью и не изменяющей своего объема. Здесь мы рассмотрим именно такую упрощенную схему. Как показывают современные экспериментальные данные (Fisher, 1971), в молодом возрасте хрусталик изотропен, что делает применяемую упрощенную схему наиболее достоверной именно для этого случая, важного для моделирования спазма аккомодации. В первом приближении допустимо не учитывать и изменение внутреннего строения хрусталика при аккомодации, а также различную эластичность ядра и коры хрусталика.
Как в состоянии покоя, так и в момент аккомодации хрусталик и его капсула находятся в состоянии динамического равновесия с цинновой связкой и внутриглазной жидкостью. Чтобы математически записать условие равновесия приложенных к капсуле хрусталика сил, необходимо знать еще величину внутреннего давления заполняющего хрусталик вещества. Это давление (П) можно назвать внутрихрусталиковым (ВХД). Хотя термин «внутрихрусталиковое давление» является новым, из уже известных фактов можно не только подтвердить существование ВХД, но и заключить, что ВХД больше, чем ВГД. Запишем условие равновесия приложенных к капсуле сил для переднего сегмента хрусталика (22) и его заднего сегмента (23):

Силу F легко выразить через параметры волокон цинновой связки. На каждое волокно действует сила ??E?, где ? — радиус волокон, Е — их модуль упругости, ? — относительная деформация при растлении. Результирующая сила F будет выражаться как nfR/?(l^2-R^2 ) где n — число волокон на данном сегменте, l — длина волокон при их растяжении.
Обобщенный закон Гука для капсулы хрусталика не будет отличаться от обобщенного закона Гука для склеры. С учетом этого можно выразить упругие напряжения в капсуле хрусталика (?1 и ?2) через модуль упругости капсулы Ек, объемы сегментов хрусталика при П — Р = 0. V0,x1 и V0,x2 приращения объемов сегментов при данном П по сравнению с объемами при П — Р = 0 (приращения обозначим через ?Vx1 и ?Vx2).
Величину S для шарового сегмента можно выразить как ?(r2 + h2), где r — радиус основания сегмента, а h — его высота. Объем сегмента равен 1/3*(?h2(3r-h)) или 1/6*(?h(3r2+h2)), а величины R и r связаны соотношением R = (r2 + h2)/2h
Рассмотрим теперь биомеханическую модель аккомодации и ее спазма.
На основании всего сказанного обобщенный закон Гука для капсулы хрусталика можно записать так:

где обозначения R10 и h10 относятся к случаю, когда П - Р = 0.
Условие равновесия приложенных к капсуле сил с учетом всех сделанных уточнений и выражений (24) и (25) можно записать следующим образом:
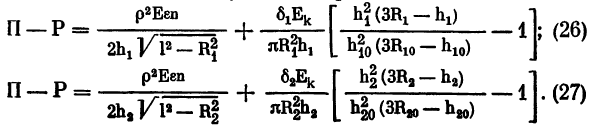
Выражения (26) и (27) верны и для состояния покоя аккомодации, для ее напряжения и спазма. Изменяются только численные значения входящих в эти выражения величин (l, R1 R2, h1, h2).
Rohen, Rentch (1969) не подтвердили наличия в цилиарном теле трех мышечных образований (мышц Мюллера, Брюкке и Иванова). Цилиарная мышца — высоко дифференцированное образование, волокна которого по своему ходу изменяют направление из продольного в радиальное, а затем циркулярное. В одной и той же мышце после инсталляции холиномиметиков волокна имеют преимущественно круговое, а после введения симпатомиметиков — продольное расположение. Кроме того, при миопической рефракции наиболее выражен продольный ход волокон, а при гиперметропической — циркулярный.
Можно заметить, что сложное строение цилиарной мышцы еще не свидетельствует о невозможности называть продольное направление волокна мышцей Брюкке, а циркулярные волокна — мышцей Мюллера. Ясно лишь, что три «мышцы» нельзя считать анатомически и функционально независимыми друг от друга.
Моделирование процесса перехода ложной миопии в осевую. Упругие напряжения в склере при шаровидной форме глаз. При эмметропии и ложной миопии упругие напряжения в склере сферического глаза определяются как ? = (P0*R)/2? [(см. формулу (1)]. Меридианальные и экваториальные (кольцевые) напряжения при этом равны ?s = ?т.
И. Ферфильфайн (1973) учел также напряжения от силы тяжести (давление вниз внутриглазной жидкости и возникающие при движениях головы инерционные напряжения). Если соответствующие меридианальные и тангенциальные напряжения обозначить через ??я, ?тт, ?sин, ?ин, то суммарное напряжение будет равно:
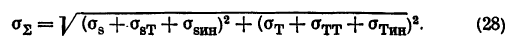
Однако автором были получены сверхвысокие значения (более 200 мм рт. ст.) эквивалентных давлений, необходимых для создания в оболочках нормальных глаз тех же напряжений, что и в увеличенных сферических глазах с истонченной склерой.
Более точная расчетная схема создана и детально исследована А. И. Дашевским и В. М. Львовским, показавшими, что в живом глазу при повышении ВГД имеет место и растяжение склеры, и сжатие сосудов.
Важную и основную роль играет повышение ВГД при конвергенции (А. И. Дашевский, 1973; С Г. Кривенков, 1973) и при переводе взгляда со строки на строку во время чтения (Л. А. Дымшиц, 1970).
Очевидно, что в точное количественное выражение для упругих напряжений склеры шаровидных глаз должны входить напряжения, вызванные повышением ВГД, силой тяжести и различными инерционными нагрузками. При расчете последних необходимо учитывать реакцию не только склеры, но и внутриглазного сосудистого ложа.
Факторы, приводящие к возникновению остаточных деформаций склеры. Как было показано выше, при ложной и осевой миопии конвергентное повышение ВГД больше, чем при эмметропии. Это связано с меньшей величиной угла начала деформации склеры наружной прямой мышцей у лиц с ложной и осевой миопией. У них конвергентное повышение ВГД начинается раньше и при равных значениях а достигает более высокого уровня, чем у лиц с нормальной остротой зрения. Упругие напряжения в мышцах при спазме усиливаются за счет увеличения модуля упругости. Можно допустить, что величина Еm различна для наружной прямой мышцы глаза при эмметропии и миопии. Конвергентное повышение ВГД служит наиболее достоверным (в настоящее время) экспериментальным доказательством наличия спазма мышц-абдукторов при ложной миопии.
Деформация склеры при конвергенции в норме является примером обратимой деформации.
Для возникновения остаточных деформаций склеры при переходе ложной миопии в осевую упругие напряжения в склере должны достичь уровня, соответствующего пределу текучести для ткани склеры.
К повышению напряжений в склере приводит совокупность факторов. Некоторые из них изменяют упругие напряжения, повышая ВГД. Они создают при зрительной работе на близком расстоянии повышенную статическую нагрузку на склеру.
Зрительная работа на близком расстоянии сопровождается не только конвергенцией, но и наклоном туловища вперед. Влияние наклоненного положения туловища на офтальмотонус здоровых глаз исследовала Т. С. Павлова (1962) с помощью тонометра Дашевского.
Обозначим через Р? величину ВГД при наклоне туловища вперед на угол ? по сравнению с вертикальным положением. Т. С. Павлова исследовала 50 человек, измерив у них Р? в положении сидя (это соответствует углу ? = 0 и ВГД: Р = Р0), в положении лежа на боку (?= 90°), Измерения проводили через 5 мин после перевода испытуемого в новое положение (табл. 10).

Кроме того, автор дополнительно исследовала 10 человек с целью выяснить влияние на офтальмотонус угла наклона головы вниз по отношению к туловищу. Обозначим эАт угол через у, Результаты специально проведенных на 10 испытуемых измерений с изменением как угла ф, так и угла у приведены в табл. 11.

Приведенные данные позволяют судить о повышении офтальмотонуса при наклоне туловища. Позже почти аналогичные данные были получены И. Л. Ферфильфайном (1973).
При реально существующем угле наклона туловища при зрительной работе на близком расстоянии повышение ВГД небольшое (до 2 мм рт. ст.) и играет, следовательно, меньшую роль, чем конвергентное. Тем не менее общее повышение ВГД при зрительной работе на близком расстоянии становится за счет наклона туловища еще более значительным. Оно может достичь 5—7 мм рт. ст., что создает ощутимую статическую нагрузку на склеру. При длительной работе такая статическая нагрузка является одной из причин снижения предела применимости закона Гука (то есть предела пропорциональности упругих напряжений в склере ее объемным деформациям).
Другая часть факторов, увеличивающих упругие напряжения в склере, делает нагрузку на ткани склеры динамической.
Роль толчкообразного характера изменений ВГД при чтении в развитии миопии подчеркивали JI. А.Дымшиц (1970), А. И. Дашевский (1973), Cornberg (1954). Такие инерционные нагрузки периодического характера сопровождают как конвергенцию, так и другие движения глаз при зрительной работе на близком расстоянии. Более правильно поэтому говорить о периодическом характере изменений упругих напряжений в склере, которые циклически возрастают по сравнению со значениями, достигнутыми за счет статической нагрузки» Инерционные нагрузки, возникающие при движениях головы, обычно также имеют периодический характер. Следовательно, и они создают динамическую нагрузку на склеру. Менее важные факторы — суточные, пульсовые колебания ВГД — по механизму своего влияния на упругие напряжения склеры не отличаются от рассмотренных. Важно понимать, что остаточные деформации не могут возникать только под влиянием изменений ВГД периодического характера без статического фона, уровень которого возрастает в основном за счет конвергентного повышения ВГД.
Суммируя все сказанное выше, можно сделать вывод о том, что остаточные деформации склеры, то есть ее необратимое растяжение, вызываются рядом факторов, проявляющихся при зрительной работе на близком расстоянии. Однако у разных лиц эти факторы могут проявляться в разной степени как в силу индивидуальных особенностей склеры и экстраокулярных мышц, так и в силу индивидуальных особенностей посадки при зрительной работе. Поскольку спазм аккомодации приводит к необходимости приблизить текст к глазам, он является главным пусковым механизмом описанных выше процессов. Тем не менее условия для перехода ложной миопии в истинную, осевую в силу перечисленных индивидуальных особенностей могут быть различными.
Особенно способствуют возникновению осевой миопии спазм экстраокулярных мышц (увеличение Еm), меньше, чем обычно, толщина и упругость склеры, неправильная посадка с резким наклоном туловища и большим, чем это диктуется спазйом аккомодации, приближением текста к глазам.
Механизм возникновения остаточных деформаций склеры. Согласно данным А. И. Дашевского и В; М. Львовского, диаграмма ?—? (где ? — относительная деформация) имеет вид, представленный на рис. 20. Модуль упругости Е3 изменяется с изменением деформации е, как это было показано на основе анализа формулы Фриденваль- да. До некоторых напряжений, соответствующих пределу пропорциональности, деформации обратимы, то есть после снятия нагрузки остаточные деформации отсутствуют. В условиях работы на близком расстоянии, в особенности при конвергенции, происходят ударные воздействия стекловидного тела на склеру. Известно, что при динамическом воздействии напряжение и деформации выше статических от той же нагрузки. Отношение динамических величин к соответствующим статическим называется динамическим коэффициентом Кдин = ?дин/?ст, или Кдин = ?дин/?ст. При ударном (импульсном) воздействии динамический коэффициент больше 1.
Во время таких неоднократно повторяющихся ударов напряжения в склере превышают предел пропорциональности склеры. В этом случае после снятия нагрузки возникают весьма малые (микроостаточные) деформации, как это видно из рис. 21. Разгрузка происходит по линии, параллельной ОА. Тангенс угла наклона линии ОА соответствует начальному модулю упругости склеры Енач. Предел пропорциональности при последующих воздействиях становится равным тому напряжению, при котором произведена предыдущая разгрузка, то есть величине ВВ.

Из рис. 21 как будто бы следует, что если при повторных ударных воздействиях напряжения не будут превышать ?1,дин остаточные деформации не должны развиваться. В действительности дело обстоит иначе. Когда напряжения в склере меньше предела пропорциональности, они определяются формулой

Если же в склере впервые возникают напряжения, большие предела пропорциональности ?пр, появляется остаточная деформация ?ост вследствие чего увеличивается радиус склеры на dR и уменьшается ее толщина на величину d?. Тогда напряжения в склере определяются формулой

Из формулы (30) следует, что в связи с увеличением радиуса склеры и уменьшением ее толщины при одном и том же давлении Р, о есть при повторном воздействии на уже имеющуюся остаточную деформацию, оболочки импульса Р, будут возникать все большие напряжения, чем в первый раз от действия такого же импульса. Следовательно, при повторных ударных импульсах напряжения будут большими, чем ВВ (рис. 22). Микродеформации, возрастая, складываются и возникает уже явная деформация задней части склеры, за счет чего прогрессирует близорукость.

В дальнейшем, исходя из хода диаграммы ? — ?, при одном и том же возрастании напряжения на d? остаточные деформации будут усиливаться и, следовательно, увеличивается близорукость.
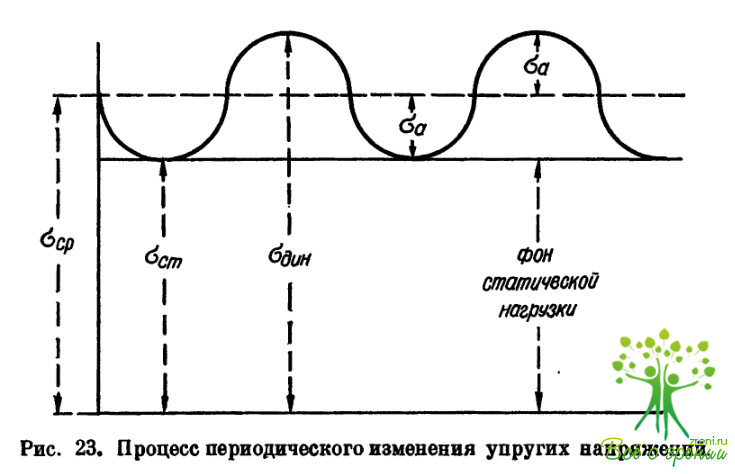
Разделение статической и динамической нагрузки позволяет также рассмотреть вопрос о роли каждой из них в достижении упругими напряжениями предела текучести. Представим изменения напряжений в виде периодического процесса (рис. 23). Как видно из рисунка, цикл награждения является знакопостоянным. Ясно, что разница ?дин — ?Ост может быть выражена как 2?а, где ?а есть ? амплитудное, отсчитываемое от ?ср = (?ст+?дин)/2. Для предельного цикла, в котором ?ст + ?дин = ?тек, при отсутствии статической нагрузки величина 2?а = ?тек должна достичь предела текучести, то есть ?тек. При отсутствии динамической нагрузки для достижения ?тек необходимо, чтобы ?ст = ?тек. Построим график, отражающий эти факты в координатах ?ст — ?а (рис. 24). Заштрихованная часть рисунка — область текучести тканей склеры. Значит, всегда можно по этому рисунку определить необходимый для достижения предела текучести уровень динамической нагрузки, то есть ?а. Такой график позволяет (при наличии экспериментальных данных) судить о роли различных факторов в переходе ложной миопии в осевую в каждом конкретном случае.

Для практических расчетов условий возникновения остаточных деформаций склеры важны полученные Э. С. Аветисовым и другими (1975) данные точных измерений ряда механических характеристик склеры, на основании которых были получены экспериментальные кривые зависимости между напряжением и деформацией.
Длительно действующая постоянная статистическая нагрузка, безусловно, имеет чисто упругостную форму. Динамическая же нагрузка (по крайней мере, часть динамически нагружающих факторов) относится к упруго- вязкому типу.
Из практики известно, что одного только наличия рассмотренных нами факторов, промежуточных между спазмом аккомодации (поздней псевдомиопией) и осевой миопией, для быстрого появления остаточных деформаций недостаточно. Они приводят к достаточному увеличению упругих напряжений лишь по истечении длительного периода времени (А. И. Дашевский, 1973).

Комментариев 0