Строение периферической нервной системы


Содержание:
Описание
В периферической нервной системе различают следующие компоненты:- Ганглии.
- Нервы.
- Нервные окончания и специализированные органы чувств.
↑ Ганглии
Ганглии представляют собой скопление нейронов, формирующих в анатомическом смысле небольшие узелки различного размера, разбросанные в различных участках тела. Различают два типа ганглиев — цереброспинальные и вегетативные. Тела нейронов спинномозговых ганглиев, как правило, округлой формы и различного размера (от 15 до 150 мкм). Ядро располагается в центре клетки и содержит четкое круглое ядрышко (рис. 1.5.1).
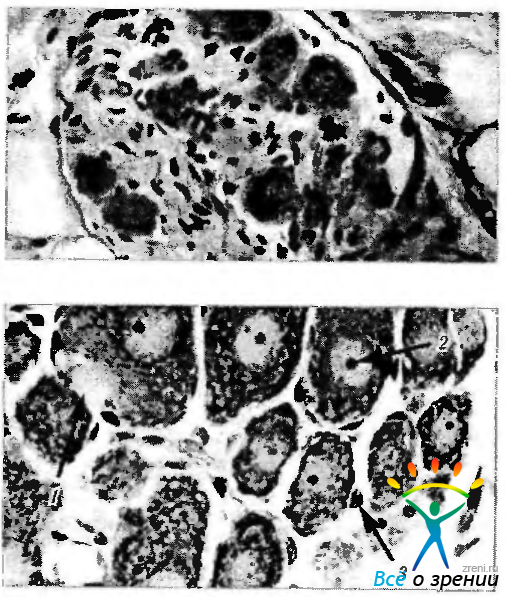
Рис. 1.5.1. Микроскопическое строение интрамурального ганглия (а) и цитологические особенности ганглиозных клеток (б): а — группы ганглиозных клеток, окруженные волокнистой соединительной тканью. Снаружи ганглий покрыт капсулой, к которой прилежит жировая клетчатка; б—нейроны ганглия (1- влючение в цитоплазме ганглиозной клетки; 2 — гипертрофированое ядрышко; 3 — клетки-сателлиты)
Каждое тело нейрона отделено от окружающей соединительной ткани прослойкой уплощенных капсулярных клеток (амфицитов). Их можно отнести к клеткам глиальной системы. Проксимальный отросток каждой ганглиозной клетки в заднем корешке разделяется на две ветви. Одна из них вливается в спинномозговой нерв, в котором проходит к рецепторному окончанию. Вторая входит в задний корешок и достигает заднего столба серого вещества на той же стороне спинного мозга.
Ганглии вегетативной нервной системы по строению сходны с цереброспинальными ганглиями. Наиболее существенное отличие сводится к тому, что нейроны вегетативных ганглиев мультиполярны. В области глазницы обнаруживаются различные вегетативные ганглии, обеспечивающие иннервацию глазного яблока.
↑ Периферические нервы
Периферические нервы являются четко определяемыми анатомическими образованиями и довольно прочны. Нервный ствол окутывается снаружи соединительнотканным футляром на всем протяжении. Этот наружный футляр называют эпинервием. Группы из нескольких пучков нервных волокон окружаются периневрием. От периневрия отделяются тяжи рыхлой волокнистой соединительной ткани, окружающие отдельные пучки нервных волокон. Это эндоневрий (рис. 1.5.2).

Рис. 1.5.2. Особенности микроскопического строения периферического нерва (продольный срез): 1— аксоны нейронов: 2— ядра шванновских клеток (леммоциты); 3—перехват Ранвье
Периферические нервы обильно снабжены кровеносными сосудами.
Периферический нерв состоит из различного количества плотно упакованных нервных волокон, являющихся цитоплазматическими отростками нейронов. Каждое периферическое нервное волокно покрыто тонким слоем цитоплазмы — неврилеммой, или шванновской оболочкой. Шванновские клетки (леммоциты), участвующие в формировании этой оболочки, происходят из клеток нервного гребня.
В некоторых нервах между нервным волокном и шванновской клеткой располагается слой миелина. Первые называются миелинизированными, а вторые — немиелинизированными нервными волокнами.
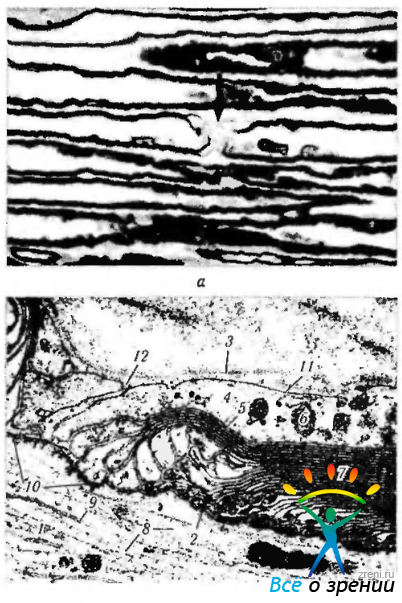
Миелин (рис. 1.5.3)
[banner_centerrs] {banner_centerrs} [/banner_centerrs]
Рис. 1.5.3. Периферический нерв. Перехваты Ранвье: а — светооптическая микроскопия. Стрелкой указан перехват Ранвье; б—ультраструктурные особенности (1—аксоплазма аксона; 2— аксолемма; 3 — базальная мембрана; 4 — цитоплазма леммоцита (шванновская клетка); 5 — цитоплазматическая мембрана леммоцита; 6 — митохондрия; 7 — миелиновая оболочка; 8 — нейрофилламенты; 9 — нейротрубочки; 10 — узелковая зона перехвата; 11 - плазмолемма леммоцита; 12 — пространство между соседними леммоцитами)
покрывает нервное волокно не сплошь, а через определенное расстояние прерывается. Участки прерывания миелина обозначаются перехватами Ранвье. Расстояние между последовательными перехватами Ранвье варьирует от 0,3 до 1,5 мм. Перехваты Ранвье имеются и в волокнах центральной нервной системы, где миелин образует олигодендроциты (см. выше). Нервные волокна разветвляются именно в перехватах Ранвье.
Каким образом формируется миелиновая оболочка периферических нервов? Первоначально шванновская клетка обхватывает аксон, так что он располагается в желобке. Затем эта клетка как бы наматывается на аксон. При этом участки цитоплазматической мембраны по краям желобка вступают в контакт друг с другом. Обе части цитоплазматической мембраны остаются соединенными, и тогда видно, что клетка продолжает обматывать аксон по спирали. Каждый виток на поперечном разрезе имеет вид кольца, состоящего из двух линий цитоплазматической мембраны. По мере наматывания цитоплазма шванновской клетки выдавливается в тело клетки.
Некоторые афферентные и вегетативные нервные волокна не имеют миелиновой оболочки. Тем не менее они защищены шванновскими клетками. Это происходит благодаря вдавливанию аксонов в тело шванновских клеток.
Механизм передачи нервного импульса в немиелинизированном волокне освещен в руководствах по физиологии. Здесь мы лишь кратко охарактеризуем основные закономерности процесса.
Известно, что цитоплазматическая мембрана нейрона поляризованна, т. е. между внутренней и наружной поверхностью мембраны существует электростатический потенциал, равный — 70 мВ. Причем внутренняя поверхность обладает отрицательным, а наружная положительным зарядом. Подобное состояние обеспечивается действием натрий-калиевого насоса и особенностями белкового состава внутрицитоплазматического содержимого (преобладание отрицательно заряженных белков). Поляризованное состояние называют потенциалом покоя.
При стимуляции клетки, т. е. нанесении раздражения цитоплазматической мембраны самыми разнообразными физическими, химическими и др. факторами, первоначально наступает деполяризация, а затем реполяризация мембраны. В физико-химическом смысле при этом наступает обратимое изменение в цитоплазме концентрации ионов К и Na. Процесс реполяризации активный с использованием энергетических запасов АТФ.
Волна деполяризации — реполяризации распространяется вдоль цитоплазматической мембраны (потенциал действия). Таким образом, передача нервного импульса есть не что иное, как распространяющаяся волна потенциала действия.
Каково же значение в передаче нервного импульса миелиновой оболочки? Выше указано, что миелин прерывается в перехватах Ранвье. Поскольку только в перехватах Ранвье цитоплазматическая мембрана нервного волокна контактирует с тканевой жидкостью, только в этих местах возможна деполяризация мембраны таким же образом, как в немиелинизированных волокнах. На остальном протяжении этот процесс невозможен в связи с изолирующими свойствами миелина. В результате этого между перехватами Ранвье (от одного участка возможной деполяризации до другого) передача нервного импульса осуществляется внутрицитоплазматическими местными токами. Поскольку электрический ток проходит гораздо быстрее, чем непрерывная волна деполяризации, передача нервного импульса в миелинизированном нервном волокне происходит значительно быстрее (в 50 раз), причем скорость увеличивается с увеличением диаметра нервного волокна, что обусловлено снижением внутреннего сопротивления. Подобный тип передачи нервного импульса называется сальтаторным. т. е. прыгающим. Исходя из изложенного, видно важное биологическое значение миелиновых оболочек.
↑ Нервные окончания
Афферентные (чувствительные) нервные окончания (рис. 1.5.5, 1.5.6).

Рис. 1.5.5. Особенности строения различных рецепторных окончаний: а — свободные нервные окончания; б— тельце Мейснера; в — колба Краузе; г — тельце Фатер-Пачини; д — тельце Руффини
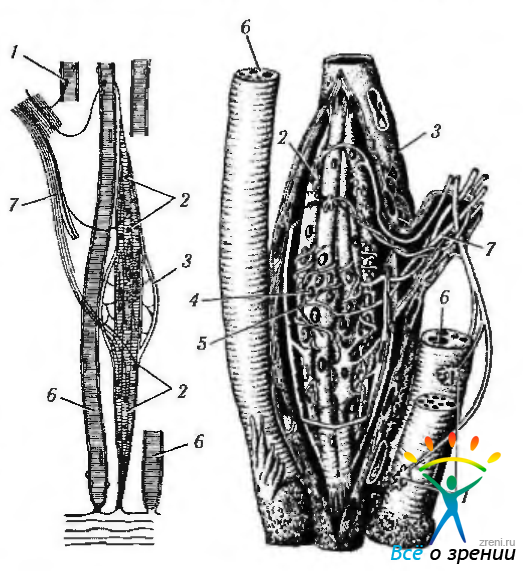
Рис. 1.5.6. Строение нервно-мышечного веретена: а—моторная иннервация интрафузальных и экстрафузальных мышечных волокон; б спиральные афферентные нервные окончания вокруг интрафузальных мышечных волокон в области ядерных сумок (1 — нервно-мышечные эффекторные окончания экстрафузальных мышечных волокон; 2 — моторные бляшки интрафузальных мышечных волокон; 3 — соединительнотканная капсула; 4 — ядерная сумка; 5 — чувствительные кольцеспиральные нервные окончания вокруг ядерных сумок; 6 — скелетные мышечные волокна; 7 — нерв)
Афферентные нервные окончания представляют собой концевые аппараты дендритов чувствительных нейронов, повсеместно располагающихся во всех органах человека и дающие информацию центральной нервной системе об их состоянии. Воспринимают они раздражения, исходящие и из внешней среды, преобразуя их в нервный импульс. Механизм возникновения нервного импульса характеризуется уже описанными явлениями поляризации и деполяризации цитоплазматической мембраны отростка нервной клетки.
Существует ряд классификаций афферентных окончаний — в зависимости от специфичности раздражения (хеморецепторы, барорецепторы, механорецепторы, терморецепторы и др.), от особенностей строения (свободные нервные окончания и несвободные).
Обонятельные, вкусовые, зрительные и слуховые рецепторы, а также рецепторы, воспринимающие движение частей тела относительно направления силы тяжести, называют специальными органами чувств. В последующих главах этой книги мы подробно остановимся только на зрительных рецепторах.
Рецепторы разнообразны по форме, строению и функциям. В данном разделе нашей задачей не является подробное описание различных рецепторов. Упомянем лишь о некоторых из них в разрезе описания основных принципов строения. При этом необходимо указать на различия свободных и несвободных нервных окончаний. Первые характеризуются тем, что они состоят только из ветвления осевых цилиндров нервного волокна и клетки глии. При этом они контактируют разветвлениями осевого цилиндра с клетками, возбуждающими их (рецепторы эпителиальных тканей). Несвободные нервные окончания отличаются тем, что в своем составе они содержат все компоненты нервного волокна. Если они покрыты соединительнотканной капсулой, они называются инкапсулированными (тельце Фатер—Пачини, осязательное тельце Мейснера, терморецепторы колбы Краузе, тельца Руффини и др.).
Разнообразно строение рецепторов мышечной ткани, часть которых обнаруживается в наружных мышцах глаза. В этой связи на них мы остановимся более подробно. Наиболее распространенным рецептором мышечной ткани является нервно-мышечное веретено (рис. 1.5.6). Это образование регистрирует растяжение волокон поперечно-полосатых мышц. Представляют они собой сложные инкапсулированные нервные окончания, обладающие как чувствительной, так и двигательной иннервацией. Число веретен в мышце зависит от ее функции и тем выше, чем более точными движениями она обладает. Нервно-мышечное веретено располагается вдоль мышечных волокон. Веретено покрыто тонкой соединительнотканной капсулой (продолжение периневрия), внутри которой находятся тонкие поперечнополосатые интрафузальные мышечные волокна двух видов:
- волокна с ядерной сумкой — в расширенной центральной части которых содержатся скопления ядер (1—4— волокна/веретено);
- волокна с ядерной цепочкой — более тон кие с расположением ядер в виде цепочки в центральной части (до 10 волокон/веретено).
Чувствительные нервные волокна образуют кольцеспиральные окончания на центральной части интрафузальных волокон обоих типов и гроздьевидные окончания у краев волокон с ядерной цепочкой.
Двигательные нервные волокна — тонкие, образуют мелкие нервно-мышечные синапсы по краям интрафузальных волокон, обеспечивая их тонус.
Рецепторами растяжения мышцы являются также нервно-сухожильные веретена (сухожильные органы Гольджи). Это веретеновидные инкапсулированные структуры длиной около 0,5—1,0 мм. Располагаются они в области соединения волокон поперечнополосатых мышц с коллагеновыми волокнами сухожилий. Каждое веретено образовано капсулой из плоских фиброцитов (продолжение периневрия), которая охватывает группу сухожильных пучков, оплетенных многочисленными терминальными веточками нервных волокон, частично покрытых леммоцитами. Возбуждение рецепторов возникает при растяжении сухожилия во время мышечного сокращения.
Эфферентные нервные окончания несут информацию от центральной нервной системы к исполнительному органу. Это окончания нервных волокон на мышечных клетках, железах и др. Более подробное их описание будет приведено в соответствующих разделах. Здесь мы подробно остановимся лишь на нервно-мышечном синапсе (моторная бляшка). Моторная бляшка располагается на волокнах поперечнополосатых мышц. Состоит она из концевого ветвления аксона, образующего пресинаптическую часть, специализированного участка на мышечном волокне, соответствующего постсинаптической части, и разделяющей их синаптической щели. В крупных мышцах один аксон иннервирует большое количество мышечных волокон, а в небольших мышцах (наружные мышцы глаза) каждое мышечное волокно или их небольшая группа иннервируется одним аксоном. Один мотонейрон в совокупности с иннервируемыми им мышечными волокнами образует двигательную единицу.
Пресинаптическая часть формируется следующим образом. Вблизи мышечного волокна аксон утрачивает миелиновую оболочку и дает несколько веточек, которые сверху покрыты уплощенными леммоцитами и базальной мембраной, переходящей с мышечного волокна. В терминалах аксона имеются митохондрии и синаптические пузырьки, содержащие ацетилхолин.
Синаптическая щель имеет ширину 50 нм. Располагается она между плазмолеммой ветвлений аксона и мышечного волокна. Содержит она материал базальной мембраны и отростки глиальных клеток, разделяющих соседние активные зоны одного окончания.
Постсинаптическая часть представлена мембраной мышечного волокна (сарколеммой), образующей многочисленные складки (вторичные синаптические щели). Эти складки увеличивают общую площадь щели и заполнены материалом, являющимся продолжением базальной мембраны. В области нервно-мышечного окончания мышечное волокно не имеет исчерченности. содержит многочисленные митохондрии, цистерны шероховатого эндоплазматического ретикулума и скопление ядер.
Механизм передачи нервного импульса на мышечное волокно сходен с таковым в химическом межнейронном синапсе. При деполяризации пресинаптической мембраны происходит выделение ацетилхолина в синаптическую щель. Связывание ацетилхолина с холинорецепторами в постсинаптической мембране вызывает ее деполяризацию и последующее сокращение мышечного волокна. Медиатор отщепляется от рецептора и быстро разрушается ацетил-холинэстеразой.
↑ Регенерация периферических нервов
При разрушении участка периферического нерва в течение недели наступает восходящая дегенерация проксимальной (ближайшей к телу нейрона) части аксона с последующим некрозом как аксона, так и шванновской оболочки. На конце аксона формируется расширение (ретракционная колба). В дистальной части волокна после его перерезки отмечается нисходящая дегенерация с полным разрушением аксона, распадом миелина и последующим фагоцитозом детрита макрофагами и глией (рис. 1.5.8).

Рис. 1.5.8. Регенерация миелинового нервного волокна: а - после перерезки нервного волокна проксимальная часть аксона (1) подвергается восходящей дегенерации, миелиновая оболочка (2) в области повреждения распадается, перикарион (3) нейрона набухает, ядро смещается к периферии, хромафильная субстанция (4) распадается; б—дистальная часть, связанная с иннервируемым органом, претерпевает нисходящую дегенерацию с полным разрушением аксона, распадом миелиновой оболочки и фагоцитозом детрита макрофагами (5) и глией; в — леммоциты (6) сохраняются и митотически делятся, формируя тяжи — ленты Бюгнера (7), соединяющиеся с аналогичными образованиями в проксимальной части волокна (тонкие стрелки). Через 4—6 недель структура и функция нейрона восстанавливается, от проксимальной части аксона дистально отрастают тонкие веточки (жирная стрелка), растущие вдоль ленты Бюгнера; г — в результате регенерации нервного волокна восстанавливается связь с органом-мишенью и регрессирует ее атрофия: д — при возникновении преграды (8) на пути регенерирующего аксона компоненты нервного волокна формируют травматическую неврому (9), которая состоит из разрастающихся веточек аксона и леммоцитов
Начало регенерации характеризуется сначала пролиферацией шванновских клеток, их передвижением вдоль распавшегося волокна с образованием клеточного тяжа, лежащего в эндоневральных трубках. Таким образом, шванновские клетки восстанавливают структурную целостность в месте разреза. Фибробласты также пролиферируют, но медленнее шванновских клеток. Указанный процесс пролиферации шванновских клеток сопровождается одновременной активацией макрофагов, которые первоначально захватывают, а затем лизируют оставшийся в результате разрушения нерва материал.
Следующий этап характеризуется прорастанием аксонов в щели, образованные шванновскими клетками, проталкиваясь от проксимального конца нерва к дистальному. При этом от ретракционной колбы в направлении дистальной части волокна начинают отрастать тонкие веточки (конусы роста). Регенерирующий аксон растет в дистальном направлении со скоростью 3—4 мм сут вдоль лент из шванновских клеток (ленты Бюгнера), которые играют направляющую роль. В последующем наступает дифференциация шванновских клеток с образованием миелина и окружающей соединительной ткани. Коллатерали и терминали аксонов восстанавливаются в течение нескольких месяцев. Регенерация нервов происходит только при условии отсутствия повреждения тела нейрона, небольшом расстоянии между поврежденными концами нерва, отсутствии между ними соединительной ткани. При возникновении преграды на пути регенерирующего аксона развивается ампутационная нейрома. Регенерация нервных волокон в центральной нервной системе отсутствует.
----
Статья из книги: Строение зрительной системы человека | Вит В. В.

Комментариев 0