Методические основы современной электроретинографии


Описание
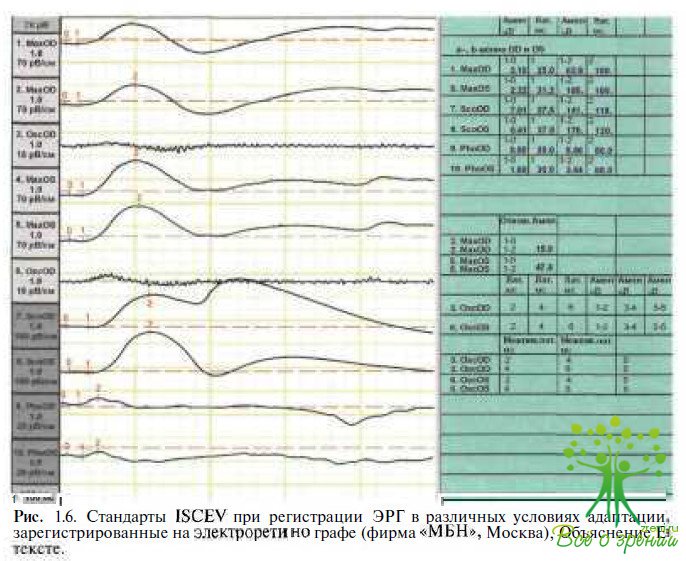
Различные типы ЭРГ отражают все многообразие структуры сетчатки, а анализ ее компонентов, генерация которых связана с активностью клеточных элементов, является диагностическим инструментом, позволяющим количественно оценить степень нарушений, локализацию, глубину и распространенность патологических процессом.Для того чтобы возможность проводить сравнительную оценку результатов электроретинографических исследований, проведенных в разных лабораториях мира, Международным обществом клинических электрофизиологов зрения (ISCEV) предложены стандарты регистрации ЭРГ, рекомендуемые для исследования зрительных функций у пациентов с различными нарушениями в зрительной системе. В клинической электроретинографии используют несколько способов регистрации ЭРГ, позволяющих выделить фотопический, скотопический и смешанный биоэлектрические ответы сетчатки. С этой целью используют соответствующие условия адаптации и стимуляции, в которых доминирует палочковая или колбочковая система сетчатки. Рекомендациями ISCEV предусмотрено проведение регистрации ЭРГ в специальных условиях световой и темновой адаптации (рис. 1.3):
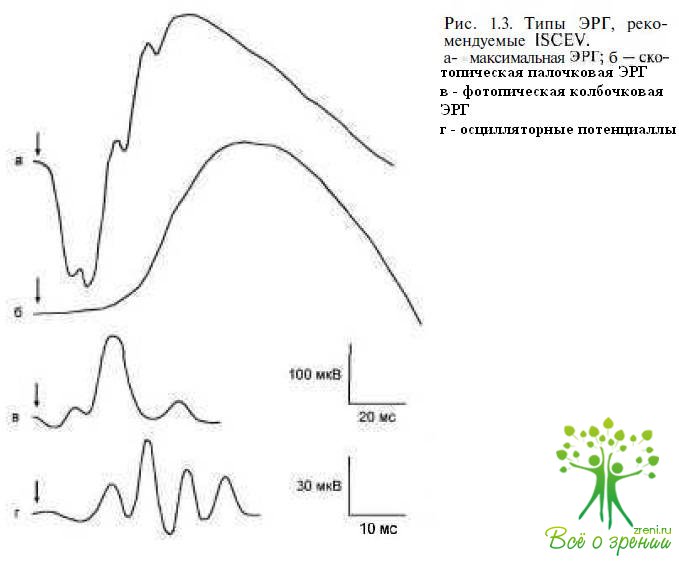
- максимальной ЭРГ в условиях темновой адаптации;
- палочковой ЭРГ в условиях темновой адаптации;
- простой колбочковой ЭРГ в условиях световой адаптации;
- осцилляторных биопотенциалов (рис. 1.4);
- фликер-ЭРГ при быстро повторяющемся стимуле 30 Гц в условиях световой адаптации (рис. 1.5).
Однако в разных лабораториях может быть использована альтернативная техника, с помощью которой должен быть записан ответ, по форме и амплитуде схожий с рекомендуемыми «Стандартами». Условия выполнения всех 5 стандартных записей могут быть изменены, возможны оригинальные условия регистрации и анализа ЭРГ.
Дополнительную информацию о функции сетчатки можно получить также с помощью других методов электроретинографического исследования, которые не входят в «Стандарты», таких как регистрация хроматической ЭРГ. макулярной ЭРГ, мультифокальной ЭРГ, ЭРГ на длительный стимул (on/off-ответ), ранних рецепторных потенциалов, регистрации порога скотопического ответа, текущей ЭРГ, ЭРГ на сдвоенные стимулы, ЭРГ, включающей S-колбочковый ответ, определение зависимости ЭРГ от интенсивности стимулирующего света (функция Naka — Rushton).
Палочковый ответ — это первый сигнал, регистрируемый после завершения темновой адаптации. Если в первые минуты темновой адаптации более чувствительна кол бочковая система (фотопическая ЭРГ), то после 15-й минуты темновой адаптации в ЭРГ преобладает биоэлектрическая активность палочковой системы (скотопическая ЭРГ). Поскольку палочки очень чувствительны к свету и дезадаптация происходит немедленно после освещения, в отличие от первого ответа на вспышку света последующие ответы имеют другую форму, латентностъ и амплитуду. Для регистрации ответа палочковой системы (скотопический ответ) ISCEV рекомендует предварительную темновую адаптацию пациента продолжительностью не менее 20 мин с целью создания стабильных условий для исследования и получения максимального скотопического ответа. Запись производят на слабую голубую или белую вспышку с энергией ниже колбочкового порога (стандартный стимул ослабляют на 2,5 лог. ед.). Минимальный интервал между стимулами 2 с. Скотопическая ЭРГ практически не имеет а-волны, а амплитуда Ь-волны в 2— 3 раза больше, чем в фотопическом ответе сетчатки.
[banner_centerrs] {banner_centerrs} [/banner_centerrs]
Максимальный, или смешанный, ответ, состоящий из комбинации ответов палочковой и колбочковой систем, регистрируют при расширенном зрачке, используя стандартные стимулы интенсивностью 1,5—4 с, с интервалом 5—10 с. ЭРГ на яркие вспышки, регистрируемая в условиях темновой адаптации, имеет большие а- и Ь-волны, чем скотопическая и фотопическая ЭРГ. И хотя в этих условиях участие колбочек достаточно велико, доминирует ответ палочковой системы. Ведущим компонентом максимального ответа является а-волна, которая отражает главным образом гиперполяризацию палочковых фоторецепторов, а склон кривой а-волны — кинетику фото-трансдукции. Позитивная Ь-волна генерируется после фоторецепторов, она связана с деполяризацией оп-биполярных клеток и клеток Мюллера.
Колбочковый, или фотопический, ответ рекомендуется регистрировать при использовании белого стандартного стимула интенсивностью 1,5— 3,0 кd x с/m2 с интервалом не менее 0,5 с в фотопических условиях на светлом фоне 17—34кd с/m2, измеренном на поверхности шара. Белый или цветной фон используют для подавления активности палочковой системы. В этих условиях колбочковый ответ достаточно стабилен и может быть воспроизведен с одними и теми же параметрами. Регистрация колбочкового ответа на единичный стимул не част усреднения множества или нескольких ответов для улучшения соотношения полезный сигнал/шум.
Колбочковую ЭРГ регистрируют после предварительной световой адаптации не менее 10 мин, так как колбочковый ответ может увеличиваться в течение этого периода. Фотопическая колбочковая ЭРГ характеризуется низкой амплитудой волн и более быстрой латентностью, чем скотопическая. Она отражает участие гиперполяризующихся оff-биполярных клеток в форме фотопической а-волны (рис. 1.6).
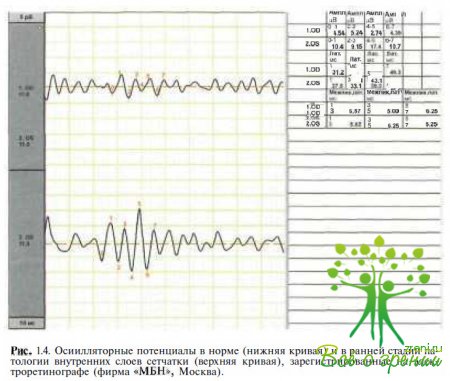
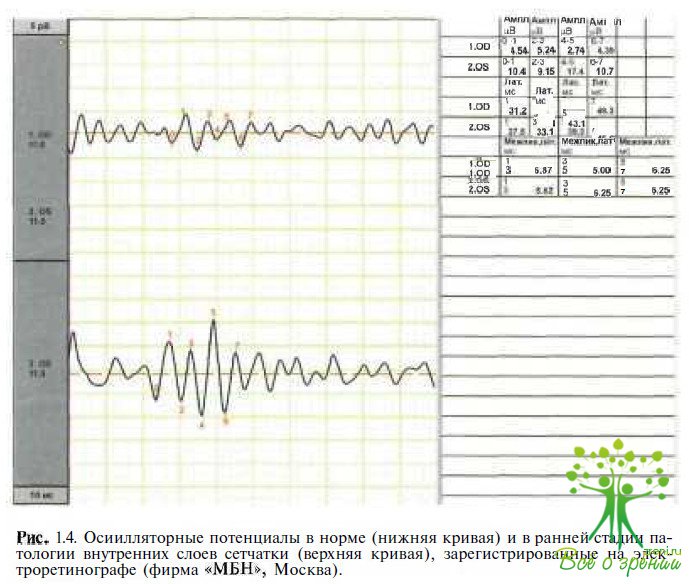
Осцилляторные биопотенциалы (ОП) представлены на ЭРГ серией волн на восходящей части Ь-волньг (5—7 вол- ночек), появляющихся после стимуляции светом большой интенсивности (см. рис. 1.4). Эти потенциалы низкой амплитуды возникают с частотой 100—160 Гц. В происхождении ОП принимают участие палочковые и колбочковые пути.
Представляя собой радиальный ток в сетчатке, ОП генерируются биполярными, амакриновыми (интерплексиформными) клетками и отражают взаимодействие клеточных элементов во внутренних слоях сетчатки. 6 зависимости от задач исследования регистрируют ОП от темно - или светоадаптированных глаз при использовании белого стандартного стимула. Полоса пропускания должна иметь нижнюю границу 75—100 Гц и верхнюю 300 Гц. После предъявления первого стимула ОП изменяются, поэтому стимулы подаются с интервалом 15 с в условиях темновой адаптации и 1,5 с при световой адаптации. Первый сигнал удаляют, сигнал, регистрируемый после предъявления второго стимула, анализируют.
Хотя ОП генерируются во внутренних слоях сетчатки, они редуцируются при патологических процессах в более Дистально расположенных отделах. Таким образом, и при макулярных дистрофиях, и при пигментном ретините возможны редуцированные, или отсутствующие, ОП. Снижение ОП отмечено при ишемических процессах в сетчатке, окклюзии вены сетчатки, диабетической ретинопатии, серповидно- клеточной ретинопатии, Х-хромосомном ювенильном ретиношизисе, стационарной ночной слепоте, болезни Бехчета и других заболеваниях.
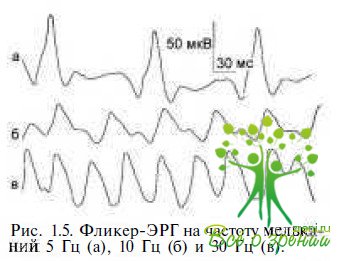
Фликер(мелькающая, ритмическая)- ЭРГ регистрируется после фотопической ЭРГ на белый стандартный стимул постоянной интенсивности с предъявляемой частотой 30 Гц, в фотопических условиях на светлом фоне, подавляющем палочковую активность. Фликерответ отражает функцию колбочковой системы сетчатки. Регистрация фликер-ЭРГ при световой адаптации уменьшает дискомфорт и позволяет стандартизировать условия фотопической адаптации. Первые ответы по форме представляют собой колбочковую ЭРГ и регистрируются в нестабильных условиях, поэтому несколько первых ответов должны быть уничтожены. Если используют нестандартные стимулы, ТО необходимы точные измерения, которые позволили бы приблизить используемый стимул к стандартным.
Выделение фотопической и скотопической активности в мелькающей ЭРГ основано на различной способности нейронов сетчатки воспроизводить световой ритм. Максимально возможная частота мельканий, воспроизводимая колбочками, — 50—100 Гц, палочками — 10—20 Гц. Для патологии палочковой системы характерно отсутствие ритмического ответа ЭРГ на низкие частоты при сохранности ответов на высокие частоты в фотопических условиях регистрации. При дистрофических изменениях в макулярной области мелькающая ЭРГ при предъявлении стимула с частотой 30 Гц не регистрируется.
Поскольку клетки Мюллера не способны реагировать на стимулы, мелькающие с частотой выше 2—4 Гц, мелькающая ЭРГ на стимулы, предъявляемые с частотой выше 4 Гц, в отличие от ЭРГ на одиночные вспышки света, является преимущественно ответом нейронов. Сравнение изменений ЭРГ на стимулы высокой и низкой (до 15 Гц) частоты в ответ на предъявление стимулов умеренной интенсивности предполагает возможность оценки степени вовлечения в патологический процесс нейронов и глиальных клеток Мюллера.
Локальная ЭРГ. Для оценки функционального состояния макулярной области разработано несколько модификаций локальной ЭРГ, регистрируемой от центральной, макулярной, области сетчатки. Однако единой методики до настоящего времени не существует. По фактической ЭРГ нельзя точно определить повреждение макулы, поскольку вклад макулярной области в кол бочковую ЭРГ невелик. Метод, разработанный в глазных болезней.
Гельмгольца, учитывает физиологические особенности сетчатки и топографию колбочковой системы. Вакуумная система обеспечивает плотное прилегание линзы-присоски к роговице, что дает возможность направлять локализацию источника света в макулярной области независимо от движения глаз и наличия центральных скотом в поле зрения, сопровождающихся нарушением фиксации взора. Источник локальной световой стимуляции (светодиод) установлен непосредственно на электроде-присоске с вмонтированной в него оптической системой, которая вместе с оптической системой глаза формирует неподвижное изображение источника света в центральной области сетчатки (рис. 1.7).

Электрод-присоска позволяет регистрировать локальную макулярную ЭРГ без контроля фиксации взора, так как надежно фиксированная на лимбе роговицы присоска передвигается синхронно с осью глаза, а источник света, закрепленный в ее центре, освещает макулярную область.
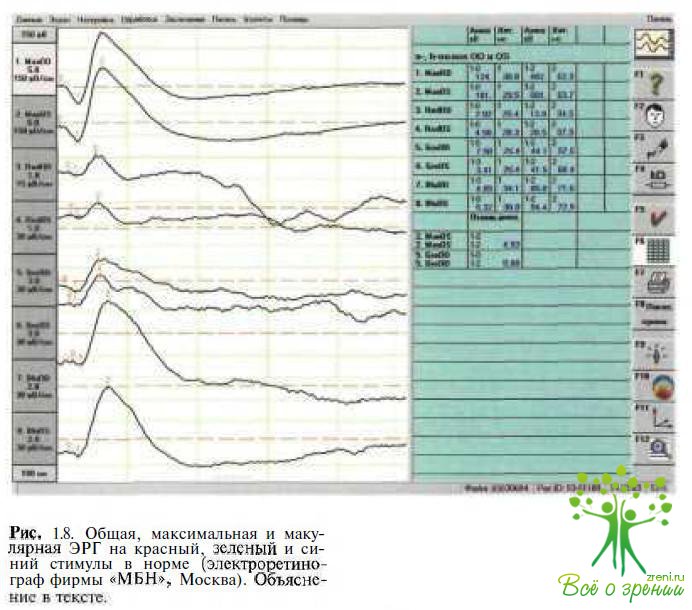
Использование красных, зеленых и синих светодиодов позволяет выделить коротковолновую (S), средневолновую (М) и длинноволновую (L) колбочковые системы, используя световой стимул разной интенсивности и длительности (рис. 1.8).
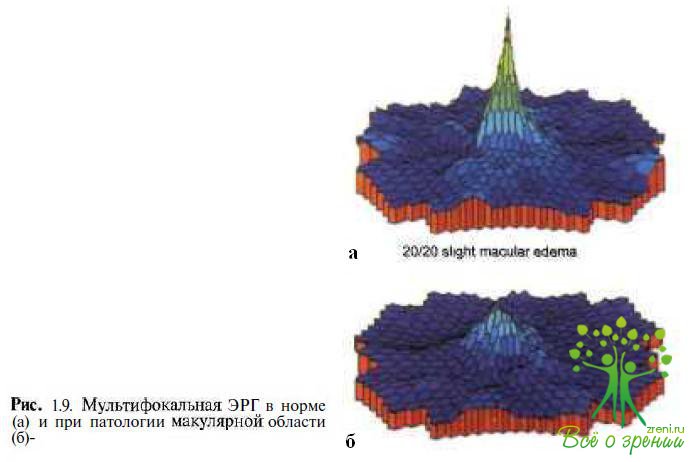
Мультифокальная ЭРГ. Мультифокальная электроретинография — метод регистрации локальных биоэлектрических ответов с каждого небольшого участка центральной области сетчатки. В качестве стимулятора используют монитор, на котором предъявляются сети гексагональных ячеек, мигающих с определенной частотой, каждая в соответствии с собственной бинарной м - последовательностью. Математическая модель представляет топографию биоэлектрической активности макулярной области в трехмерном изображении (рис. 1.9).
Различие в плотности фоторецепторов в сетчатке, отсутствие возможности контроля фиксации взора при локализации поражения в фовеальных и парафовеальных областях и наличие эксцентричной фиксации затрудняют регистрацию ответа от дакулярой области. Применение в качестве стимулятора сканирующего лазерного офтальмоскопа позволяет осуществлять одновременный контроль фиксации взора, что делает мультифокальную электроретинографию более эффективным методом исследования топографии биоэлектрической активности макулярной области сетчатки. Результаты мультифокальной электроретинографии представляются в виде ЭРГ в каждом из гексагональных участков. ЭРГ в каждом участке состоит из негативного отклонения N1, следующего за ним позитивного отклонения Р1 и второго негативного отклонения N2. Мультифокальная ЭРГ отражает функцию всех нейронов сетчатки и топографию биоэлектрической активности макулярной области, так как ее изменения наблюдаются при локализации процесса от хоровдеи до ганглиозных клеток.
Паттерн-ЭРГ. Паттерн-ЭРГ является ответом центральной сетчатки на изолюминантный стимул, который представляет собой черно-белое шахматное поле. Паттерн-ЭРГ (рис. 1.10)
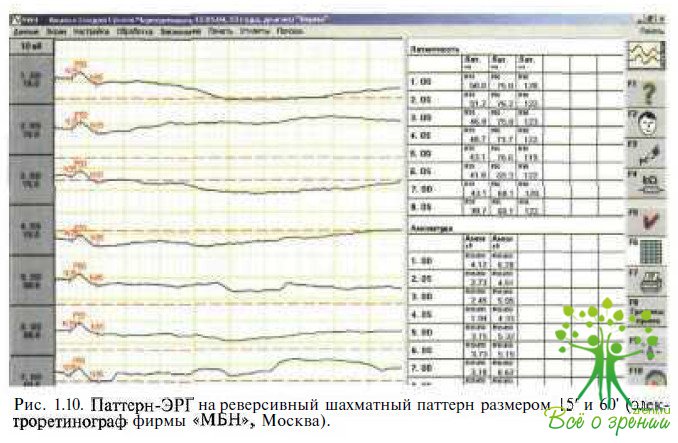
имеет два главных компонента — позитивный с амплитудой около 50 мс (Р50) и большой негативный — около 95 мс (N95). Паттерн- ЭРГ, отражающая преимущественно активность ганглиозных клеток сетчатки и функциональное состояние макулярной области, изменяется при дисфункции макулярной области, первичных и вторичных поражениях зрительного нерва. Генерируясь во внутренних слоях сетчатки, компонент Р50 паттерн-ЭРГ зависит от нормального функционирования макулярной области. Селективное отсутствие N95 отмечено при заболеваниях зрительного нерва разного происхождения. Паттерн-ЭРГ в сочетании с патгерн-ЗВП помогает при диагаостике и определении локализации патологического процесса в зрительном нерве.
Величина паттерн-ЭРГ в норме очень мала (0,5—8 мкВ) и зависит от характеристики стимула, поэтому к ее регистрации предъявляют более высокие требования, чем к записи максимальной ЭРГ. Паттерн-ЭРГ на паттернреверсивный стимул зависит от частоты предъявления и углового размера стимула, величины шахматных квадратов. Амплитуду Р50 измеряют от пика N35 до вершины волны Р50, при отсутствии N35 — от изолинии, амплитуду N95, которая включает амплитуду Р50, — от пика Р50 до изгиба или пика N95, латентность основных компонентов паттерн-ЭРГ — от стимула до пиков ее компонентов.
При величине поля стимуляции 16—30° используют стимулы трех размеров — 60', 30' и 15' при постоянной яркости стимула 80 кд/м2. Для transi- ent-паттерн-ЭРГ реверсивная частота 2—6 реверсий в 1 с (1—3 Гц), полоса пропускания усилителя 1 — 100 Гц. Паттерн-ЭРГ регистрируют без расширения зрачка с целью сохранения аккомодации и достижения наилучшего изображения паттерна на сетчатке. Для записи паттерн-ЭРГ используют роговичные электроды, не искажающие стимул, или конъюнктивальные, которые можно помещать на роговицу или в нижний свод конъюнктивы.
Зависимость ЭРГ от интенсивности стимула. Применение световых стимулов разной интенсивности связано с тем, что при низкой интенсивности в формировании ЭРГ принимают участие преимущественно палочковые компоненты сетчатки, при высокой — колбочковые, при средней интенсивности — и колбочковые, и палочковые компоненты, причем последние доминируют; при низкой интенсивности стимула а-волна отсутствует (рис. 1.11).
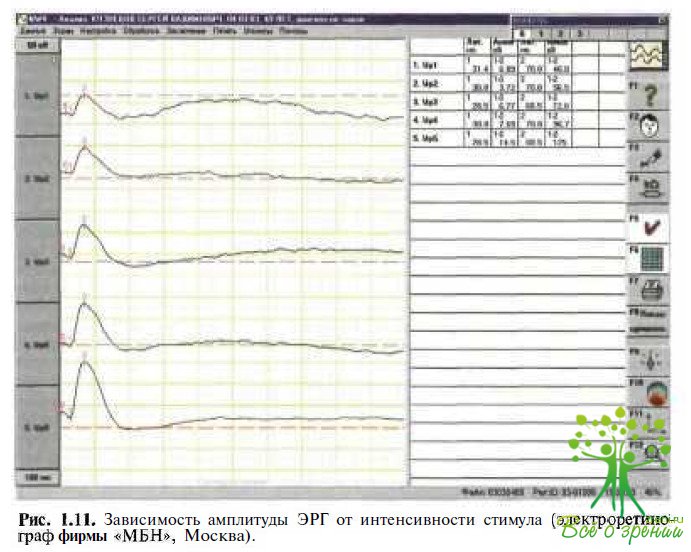
При увеличении интенсивности световых стимулов амплитуда а- и b-волн возрастает, латентность и время до пика волн (implicit time) уменьшаются. При высоких интенсивностях световых стимулов амплитуда волн достигает своего максимума (насыщения) и ее увеличение прекращается, кривая зависимости амплитуды от интенсивности стимулов выходит на «плато».
Поскольку при передаче сигнала от фоторецепторов к нейронам внутренних слоев происходит магнификация электрической активности от наружных слоев сетчатки к внутренним, b-волна появляется уже при интенсивности 1 лог.ед., т.е. она измеряется при низких интенсивностях, когда а-волна еще отсутствует. Таким образом, не расходуя время на длительную темновую адаптацию, в непродолжительном исследовании можно получить представление о функции колбочковой и палочковой систем, а также о чувствительности сетчатой оболочки. Это исследование целесообразно проводить у пациентов с патологией сетчатки при расчете отношения скотопической b-волны к интенсивности ганцфельдстимула. При этом определяют функцию Naka — Rush ton — соотношение стимул/ответ (S/R), которая представляет собой увеличение амплитуды b-волны ЭРГ при увеличении интенсивности стимула на 2,5 лог.ед. В норме соотношение S/R может быть представлено в виде формулы:
R/Rmax=?n(In+a)
где R — амплитуда b-волны, вызванной вспышкой интенсивностью I, представленной в футламбертах или канделах на 1 м2; ?n— интенсивность светового стимула, при которой регистрируется половина максимального ответа Rmax; a — индекс чувствительности сетчатки; экспонента п имеет величину около 1 для нормальной сетчатки и зависит от формы наклона функции S/R. Определение соотношения S/R позволяет получить информацию, которую используют для изучения происхождения различных патологических процессов в сетчатке. Так, полная потеря фоторецепторов на изолированном участке сетчатки может привести к редукции Rmax, но не влияет на величину ? или n, а снижение концентрации родопсина при патологическом укорочении наружных сегментов фоторецепторов не вызывает изменения Rmax, но способствует снижению чувствительности сетчатки.
ЭР1 на длительный стимул. Установлено, что использование стимула длительностью до 150—500 мс для стимуляции сетчатки при регистрации ЭРГ позволяет выделить в ЭРГ функцию деполяризующихся и гиперполяризующихся под воздействием света биполярных клеток (рис. 1.12).
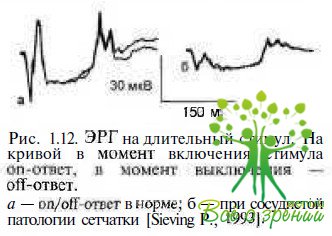
Каждая из колбочек центральной зоны сетчатки связана с двумя биполярными клетками разных типов. Деполяризующиеся биполяры (ДПБ), или оп-биполяры, дают начало on-каналу зрительной системы, который воспринимает стимулы светлее фона и реагирует на включение светового сигнала; гипсрполяризующиеся биполяры (ГПБ), или off-билоляры, воспринимают стимулы темнее фона и реагируют на выключение света. Колбочковые on- и off-бигголяры контактируют с ганглиозными клетками с оп- и off-центрами соответственно. Исследование ЭРГ на длительный белый стимул у больных с дистрофическими изменениями палочковой и колбочковой систем сетчатки позволило выделить on- b Off-ответы колбочковой (фотопической) системы и оп-ответ палочковой (скотопической). Отсутствие off-ответа при регистрации скотопической ЭРГ в норме подтверждало отсутствие off-би-поляров в палочковых путях.
Скотопический порог ответа (STR) — негативный ответ низкой амплитуды, который может быть записан после длительного периода темновой адаптации при использовании очень слабых стимулов, неспособных вызвать сигнал кол- бочковых путей или палочковую Ь-вол- ну. STR является результатом активности амакриновых клеток, которые выделяют калий в ответ на световую стимуляцию и приводят в состояние деполяризации клетки Мюллера. Клиническое значение STR пока не совсем ясно, но его отсутствие отмечено при ретиношизисе, стационарной ночной слепоте, глаукоме, вовлечении в патологический процесс клеток Мюллера.
Фотопический пороговый ответ (PTR) — корнеонегативный потенциал, регистрируемый от колбочек в фотопических условиях, который генерируется ганглиозными клетками, их аксонами и, возможно, амакриновыми клетками.
Статья из книги: Зрительные функции и их коррекция у детей | С.Э. Аветисов, Т.П. Кащенко, А.М. Шамшинова.

Комментариев 0