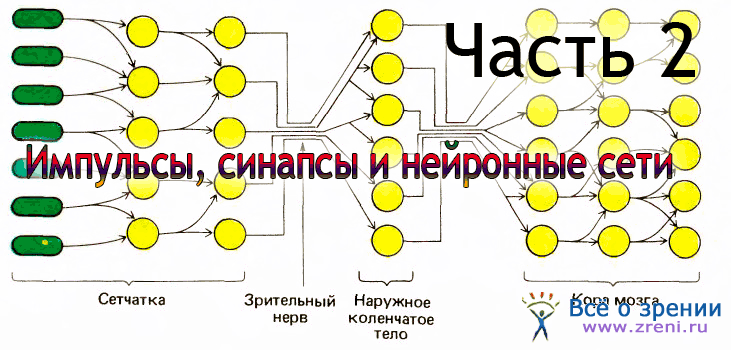
Дыхание – основа существования любого живого организма. При дыхании тело поглощает кислород из окружающей среды и выводит из организма углекислый газ – «отработанное топливо» биохимических реакций. Кислород мы расходуем практически для всех процессов, происходящих внутри нас. При этом хранить его не получается – если не дышать, запасов важного газа хватает максимум минут на пять, а дальше начинают гибнуть клетки. В состоянии покоя человек за минуту потребляет 250 мл кислорода и выдыхает 200 мл углекислого газа.