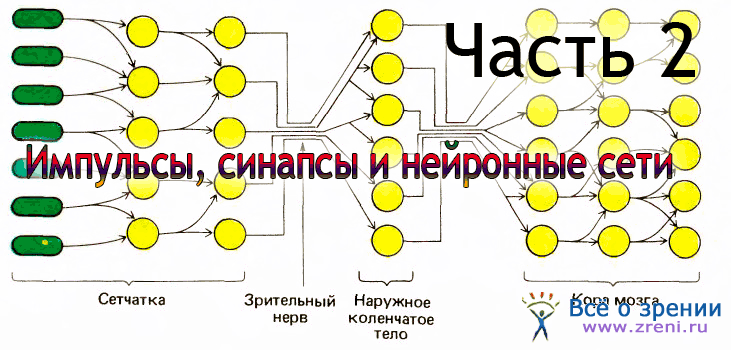
Глаз часто сравнивают с фотоаппаратом. Более уместно было бы сравнить его с телевизионной камерой, установленной на треноге, с автоматической системой слежения — машиной, которая самофокусируется, автоматически подстраивается к интенсивности света, имеет самоочищающуюся линзу и присоединена к компьютеру со столь развитыми возможностями параллельной обработки информации, что инженеры еще только начинают обсуждать сходные стратегии для конструируемой ими аппаратуры.
Гигантская работа по преобразованию света, падающего на две сетчатки, в осмысленную зрительную сцену часто странным образом игнорируется, как будто все необходимое нам для того, чтобы видеть, — это изображение внешнего мира, четко сфокусированное на сетчатке. Хотя получение резких изображений и важная задача, она скромна по сравнению с работой нервной системы — сетчатки и мозга. Как мы увидим в этой главе, вклад сетчатки уже сам по себе впечатляет.
Преобразуя свет в нервные сигналы, она начинает извлекать из окружающей среды то, что полезно, и отбрасывать то, что излишне. Никакое человеческое изобретение, включая управляемые компьютером камеры, пока еще не может соперничать с глазом. Эта глава посвящена главным образом нейронной части глаза — сетчатке, но я начну с краткого описания глазного яблока, аппарата, который содержит сетчатку и создает на ней четкое изображение внешнего мира.