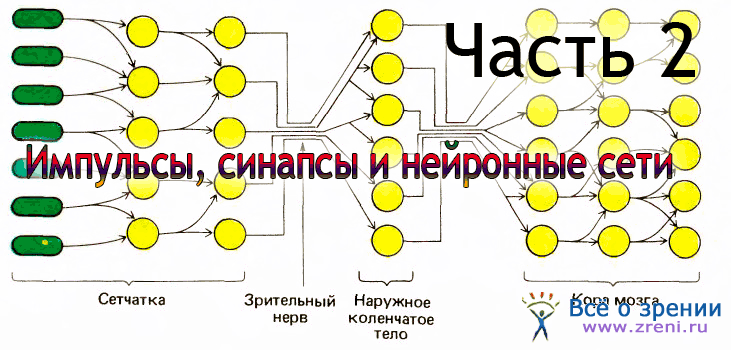
Значительную часть нейробиологии составляют разделы о том, как работают отдельные нейроны и как информация передается от клетки к клетке через синапсы. Должно быть очевидным, что без этих сведений мы окажемся в положении человека, желающего понять работу радиоприемника или телевизора, но ничего не знающего о резисторах, конденсаторах и транзисторах. За последние десятилетия благодаря изобретательности ряда нейрофизиологов, из которых наиболее известны Эндрю Хаксли, Алан Ходжкин, Бернард Катц, Джон Экклз и Стивен Куффлер, были хорошо изучены физико-химические механизмы проведения нервных импульсов и синаптической передачи. Однако столь же очевидно, что сведения такого рода сами по себе еще не могут привести к пониманию работы мозга, подобно тому как одни лишь сведения о резисторах, конденсаторах и транзисторах не позволят понять работу радиоприемника или телевизора, а знание химии чернил — прочитать пьесу Шекспира.